
| Magyarosaurus was a small sauropod from the late Cretaceous measuring only 5-6 m in length and was assigned to Titanosaurus, as T. dacus by Nopcsa in 1915. It was assigned to the new genus Magyarosaurus by Huene in 1932 in his review of saurischian dinosaurs. When compared to its relatives, the 7 m long Saltasaurus and the 20-25 m long Argentinosaurus, it’s clearly a lot smaller. Jianu and Weishampel (1999) found the humerus of Magyarosaurus was similar to that of subadults of other taxa, which suggested that dwarfing was achieved by paedomorphosis.
As an adult Magyrosaurus would have been only half the length of its mainland relative, there would have been a corresponding reduction in its body mass to one eighth. The same is seen in Telmatosaurus. All the dinosaurs in the Haţeg Region generally occupy the basal positions of their respective cladograms. They all existed during the Maastrichtian period of the late Cretaceous and they are “living fossils” in their day as they were more primitive than their contemporaries. Magyrosaurus dacus currently sits basally in the “Rapetosaurus clade” which includes a mix of early and late Cretaceous taxa. |
 Order: Saurischia Suborder: Sauropodomorpha Infraorder: Sauropoda (unranked): Titanosauria Length: 5-6 metres Late Cretaceous |
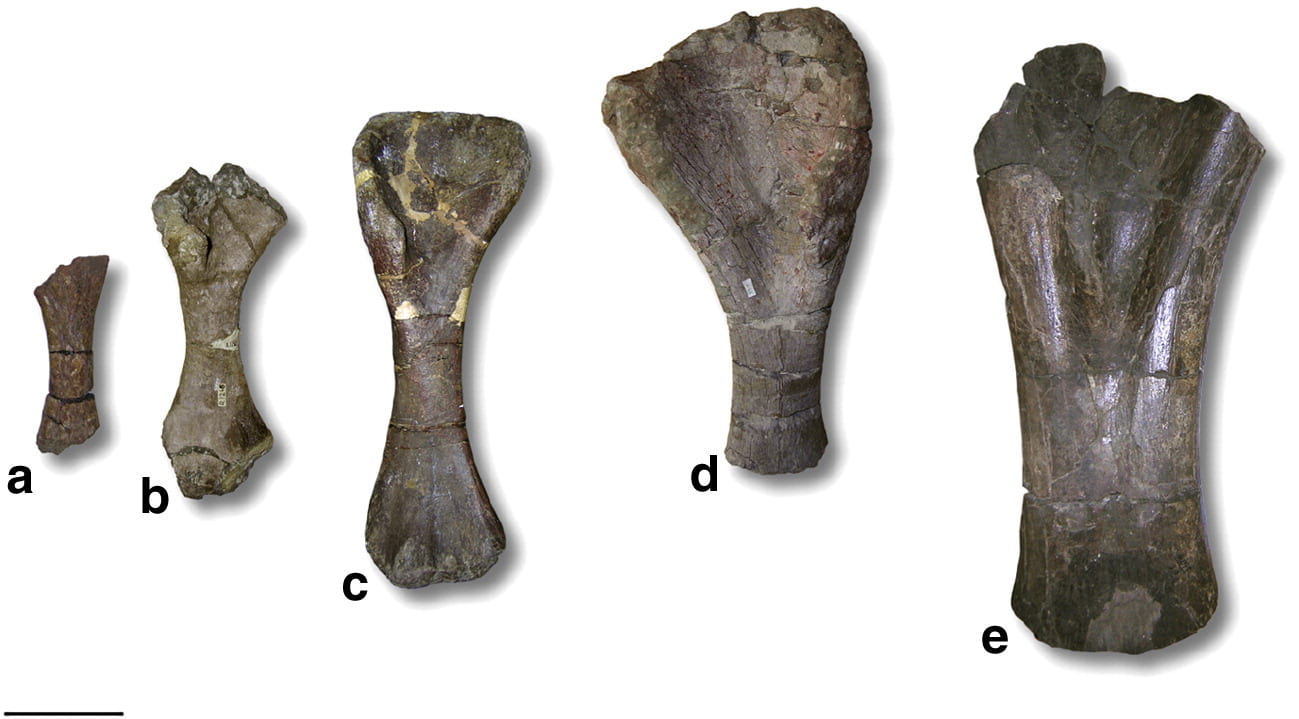 Figure 1: Photographs of some of the sampled titanosaur bones from the Maastrichtian of Romania. (a-d) Magyarosaurus dacus humeri, specimens: (a) MAFI Ob. 3092 (the smallest recorded body size, 45% maximum size); (b) FGGUB R.1246 (65% maximum size); (c) MAFI v.13492 (76% maximum size); (d) FGGUB R.1048 (largest known specimen). (e) Titanosaur indet., MAFI Ob.3104. Scale bar equals 100 mm.
Figure 1: Photographs of some of the sampled titanosaur bones from the Maastrichtian of Romania. (a-d) Magyarosaurus dacus humeri, specimens: (a) MAFI Ob. 3092 (the smallest recorded body size, 45% maximum size); (b) FGGUB R.1246 (65% maximum size); (c) MAFI v.13492 (76% maximum size); (d) FGGUB R.1048 (largest known specimen). (e) Titanosaur indet., MAFI Ob.3104. Scale bar equals 100 mm.
An ontogenetic series of Magyrosaurus dacus long bones (Figure 1) shows a histology that is only seen in older, larger members of the Sauropoda. The cortex of even the smallest specimen, which is only 45% of its maximum size (Figure 1a), is dominated by secondary osteons, where the bone has been “recycled” as part of the process of secondary remodelling, leaving only a few areas of lamellar primary fibrolamellar bone with a strong lamellar component in the bone matrix (Figure 2 a-d).
 Figure 2: Long bone histology of Magyarosaurus dacus compared with other vertebrates. (a) overview of cross section (specimen MAFI V.13492, 76% max size); (b) close-up of (a): cortex dominated by secondary remodelling. (c) Close-up of largely interstitial laminar primary bone in outermost cortex of the smallest available specimen of M. dacus (MAFI Ob.3092, 46% max size). The vascular canals are oriented circumferentially as in laminar fibrolamellar bone, but the bone matrix between the vascular canals consists largely of parallel-fibered and lamellar bone, with only a minute fraction of fibrous (or woven) bone tissue. (d) Close-up of (c). (e) Laminar fibrolamellar bone of Apatosaurus. (f) Alligator long bone histology showing lamellar-zonal bone. All scale bars equal 200 µm.
Figure 2: Long bone histology of Magyarosaurus dacus compared with other vertebrates. (a) overview of cross section (specimen MAFI V.13492, 76% max size); (b) close-up of (a): cortex dominated by secondary remodelling. (c) Close-up of largely interstitial laminar primary bone in outermost cortex of the smallest available specimen of M. dacus (MAFI Ob.3092, 46% max size). The vascular canals are oriented circumferentially as in laminar fibrolamellar bone, but the bone matrix between the vascular canals consists largely of parallel-fibered and lamellar bone, with only a minute fraction of fibrous (or woven) bone tissue. (d) Close-up of (c). (e) Laminar fibrolamellar bone of Apatosaurus. (f) Alligator long bone histology showing lamellar-zonal bone. All scale bars equal 200 µm.
A fully grown adult is indicated by the presence of an external fundamental system (Introduction: Figure 1c) in the outer bone cortex. Although this was not observed in any Magyarosaurus specimen, there was advanced secondary remodelling which is typical of the oldest histological ontogenetic stages of larger sauropods (Klein and Sander 2008). A completely remodelled long bone cortex is only found in the oldest and largest examples of Apatosaurus and Supersaurus, and this suggests that Magyrosaurus was indeed a small adult and not a juvenile.
Growth Rate in Magyrosaurus dacus
Secondary bone remodelling completely “destroyed” the primary growth record in the long bones of M. dacus without any LAGs or other growth marks making it difficult to accurately estimate the growth rate of Magyrosaurus. The strong lamellar component of the remaining primary bone, when compared with the bones of larger sauropods, suggests that the growth rate of M. dacus was much slower than its larger counterparts (Figure 2e) but still faster than endothermic reptiles that have a lamellar-zonal bone dominated cortex (Figure 2f).
 Artwork: Magyarosaurus dacus
Artwork: Magyarosaurus dacus
Kindly provided by Mihai Dumbrava
Literature cited
- Jianu, C.-M., Weishampel, D.B., 1999. The smallest of the largest: a new look at possible dwarfing in sauropod dinosaurs. Geologie en Mijnbouw 78, 335-343.
- Klein, N., Sander, M., 2008. Ontogenetic stages in the long bone histology of sauropod dinosaurs. Paleobiology 34, 248-264.
- Stein, K., Csiki, Z., Curry Rogers, K., Weishampel, D.B., Redelstorff, R., and Sander, P.M. 2010. Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria). Proceedings of the National Academy of Sciences, USA 107, 9258–9263.