
The dwarf dinosaurs of Haţeg
In 1914 the palaeontologist Franz Nopcsa suggested that Cretaceous dinosaurs of the Haţeg area of Romania showed signs of being affected by the island rule, and he observed that at the same time “while turtles, crocodilians and similar animals of the cretaceous reached their normal size, the dinosaurs almost always remain below their normal size.” The concept of the island rule is that large animals tend to diminish in scale when isolated while smaller animals in the same environment become larger. A modern example of this would be the pygmy hippopotamus (Choeropsis liberiensis) which is found in the forests and swamps of native Africa and the Millheela region in Kenya. Though examples of the island rule has been seen throughout the Pliocene, Pleistocene and Holocene in mammals of the Mediterranean islands, the “dwarf dinosaurs” at Haţeg had never been fully investigated.
Benton et al. (2010), Csiki & Benton (2010), and Weishampel et al. (2010) have re-examined data from the Haţeg area using geological evidence and dinosaur bone histology to show that Haţeg was an island in the late Cretaceous and that three species of dinosaur, the sauropod Magyarosaurus, the ornithopod Telmatosaurus and the ornithopod Zalmoxes showed signs of progenesis, a form of paedomorphosis.
Dinosaur growth rates and LAGs
How dinosaurs became so large raises interesting questions about their growth rates. Did dinosaurs have low metabolic rates and take decades to reach their full size? Or did they have high metabolic rates and reach maturity faster? Work conducted by Horner et al (1999), Sander et al. (2000), Erickson et al. (2001) and Padian et al. (2001) suggests the latter. Studies of bone histology show that dinosaurs grew according to an S-shaped (sigmoid) curve, showing a relatively slow growth early on, then rapid growth midway in development, before slowing down as the dinosaur became sexually mature and reached adult size.
The age of a dinosaur can be estimated by counting the lines of arrested growth (LAGs) as they reflect annual slow-downs in growth. It is useful to think of LAGs as the lizard/dinosaur equivalent of tree rings. The distance between each LAG indicates how much growth has occurred that year. So in the dinosaurs, in early years when the growth rate is slow the LAGs are relatively close together, and as the growth rate increases so does the gap between LAGs. As the dinosaur hits maturity the gap between LAGs become smaller again. LAGs are best seen in the fibrolamellar primary bone which indicates fast growth. As the animal grows older and matures the lamellar-zonal bone was laid down in the outer cortex, followed by denser bone with narrowly spaced LAGs in the outermost cortex.
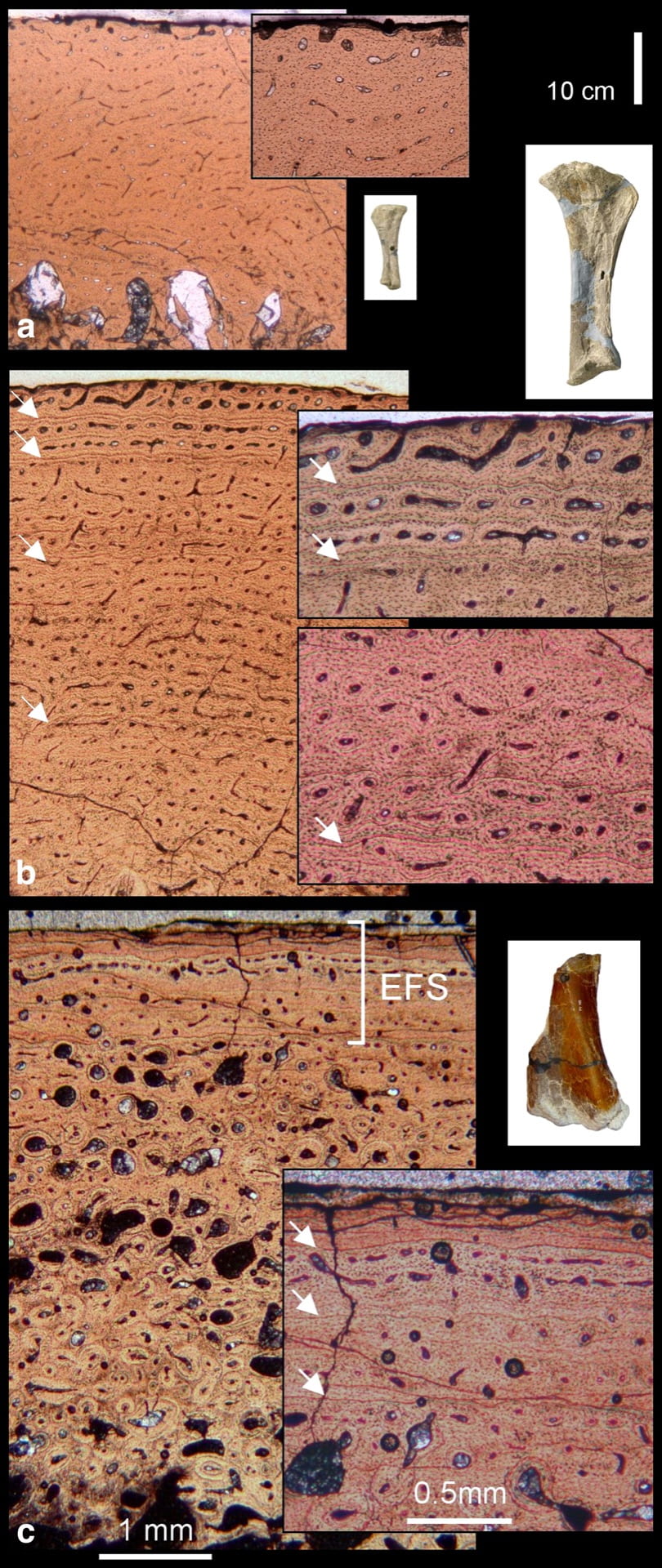 Figure 1. Histological growth series and sampled bones of Europasaurus holgeri (Mateus, Laven, and Knötsche, 2006): (a) Tibia from the smallest individual (DFMMh/FV009; body length 1.75 m). The reticular fibrolamellar tissue, which grades into laminar fibrolamellar tissue (inset), and the absence of growth marks indicate its juvenile status. (b) Tibia from a mid-sized individual (DFMMh/FV 459.5; body length 3.7 m). The cortex consists of laminar fibrolamellar bone interrupted by growth marks (arrows). Wide vascular canals opening to the outer bone surface (inset) indicate that this animal was still actively growing. (c) Distal femur from the largest individual (DFMMh/FV 415; body length 6.2 m). The external fundamental system (ESF; inset) indicates that it was fully grown. Bone surface is at the top of all photomicrographs. Black arrows indicate sample locations; white arrows indicate growth marks. Image from Sander et al. (2006).
Figure 1. Histological growth series and sampled bones of Europasaurus holgeri (Mateus, Laven, and Knötsche, 2006): (a) Tibia from the smallest individual (DFMMh/FV009; body length 1.75 m). The reticular fibrolamellar tissue, which grades into laminar fibrolamellar tissue (inset), and the absence of growth marks indicate its juvenile status. (b) Tibia from a mid-sized individual (DFMMh/FV 459.5; body length 3.7 m). The cortex consists of laminar fibrolamellar bone interrupted by growth marks (arrows). Wide vascular canals opening to the outer bone surface (inset) indicate that this animal was still actively growing. (c) Distal femur from the largest individual (DFMMh/FV 415; body length 6.2 m). The external fundamental system (ESF; inset) indicates that it was fully grown. Bone surface is at the top of all photomicrographs. Black arrows indicate sample locations; white arrows indicate growth marks. Image from Sander et al. (2006).
An island dwarf should be smaller than it closest mainland relative. One of the best examples of insular dwarfism among dinosaurs is Europasaurus from the Kimmeridgian of northern Germany (Sander et al. 2006). Individuals showing a range of bone histology from juveniles to adults (Figure 1) were found in the area and ranged in length from 1.7 to 6.2 m. At 6.2 m long, and adult is one-third shorter than its closest relative Camarasaurus. The largest Europasaurus showed histological charactistics of adulthood:
- the inner cortical fibrolamellar bone is extensively remodelled by secondary osteons that nearly obliterate the primary bone;
- the outer cortex has closely spaced LAGs inducating radical slow-down in growth rate;
- the outer zone shows characteristics of being an external fundamental system (outer cortex is avascular and consists of lamellar bone).
Literature cited
- Benton, M.J., Csiki, Z., Grigorescu, D., Redelstorff, R., Sander, P.M., Stein, K., and Weishampel, D.B. 2010. Dinosaurs and the island rule: The dwarfed dinosaurs from Hațeg Island. Palaeogeography, Palaeoclimatology, Palaeoecology 293, 438-454. pdf.
- Csiki, Z. and Benton, M.J. 2010. An island of dwarfs – reconstructing the Late Cretaceous Hațeg palaeoecosystem. Palaeogeography, Palaeoclimatology, Palaeoecology 293, 265-270. pdf.
- Erickson, G.M., Curry Rogers, K., and Yerby, S.A. 2001. Dinosaurian growth patterns and rapid avian growth rates. Nature 412, 429-433.
- Horner, J.R., de Ricqlès, A., and Padian, K. 1999. Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology. Paleobiology 25, 295-304.
- Padian, K., de Ricqlès, A., Horner, J.R. 2001. Dinosaurian growth rates and bird origins. Nature 412, 405-408.
- Sander, P.M. 2000. Long bone histology of the Tendaguru sauropods: implications for growth and biology. Paleobiology 26, 466-488.
- Sander, P. M., Mateus, O., Laven, T., and Knötschke, N. 2006. Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature 441, 739–741.
- Weishampel, D.B., Csiki, Z., Benton, M.J., Grigorescu, D., and Codrea, V. 2010. Palaeobiogeographic relationships of the Hațeg biota – Between isolation and innovation. Palaeogeography, Palaeoclimatology, Palaeoecology 293, 419-427. pdf.